
Lac Operon in E. Coli- Principle of transcriptional regulation
Explore the lac operon of Escherichia coli, a critical component in lactose fermentation. Learn how the lac repressor protein regulates gene expression and the role of primary and secondary operators in this essential biological process. Discover more about how E. coli utilizes lactose for energy.
STUDY ZONE
Operon Model
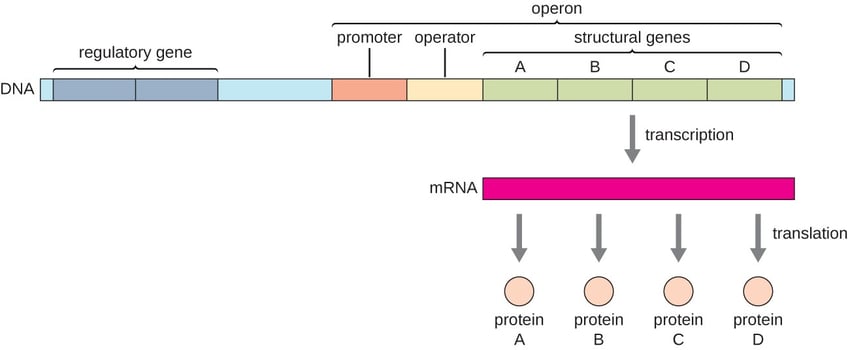
The classical operon model formulated by Jacob and Monod in 1961 provides a basic description of the gene regulation at the transcriptional level in bacteria. An operon is a functional unit of bacterial gene expression containing structural genes, a promoter, and cis-acting regulatory elements recognized by a regulatory gene product. All the structural genes in an operon are transcribed into a single mRNA, again a very common arrangement within prokaryotic genomes. The promoter of an operon serves as the initiation site for transcription of all the structural genes controlling the rate of their synthesis. The strength of the promoter is critical and determines the basal level of gene transcription, whether through a weak or a strong promoter.
Not all proteins a cell can make must be present at all times or in the same amounts, however, in bacteria. For example, enzymes to break down sugars such as lactose do not have to be produced if lactose is unavailable or not present in the surrounding cell environment. When a certain metabolite is not present, the enzymes to break it down are also not required, and to synthesize these would serve no useful purpose as well as waste the building blocks. In an inducible system, enzymes responsible for the breakdown of a specific compound will be produced by the cell only if that compound is present. The best example of an inducible system is the catabolic breakdown in E. coli of the disaccharide lactose.
Conversely, for an anabolic process, for example, the synthesis of tryptophan, enzymes are needed. But if there is plenty of tryptophan in the cell, then it does not need enzymes for its synthesis. If the enzymes for synthesizing a particular substance are produced by the cell only when that substance is absent, then the system can be termed repressible. The best known system of all those studied so far which is repressible is that of tryptophan biosynthesis in E. coli.
Genes coding for enzymes catalyzing catabolic pathways typically are expressed only when present with their substrates. The expression of these genes is induced. Typically, the product of the process represses genes encoding enzymes for anabolic pathways.
The Lac Operon of Escherichia coli
Escherichia coli (E. coli) is one of the bacteria that can ferment lactose, milk sugar, for energy and carbon sources. When this sugar is present in the environment, the lac operon, comprising an operator and three structural genes, degrades it into galactose and glucose.
The operator is a region in DNA that binds with the lac repressor protein to regulate the expression of genes located in an operon. The repressor prevents RNA polymerase from initiating transcription at the promoter if it is bound to the operator and thereby prevents gene expression.
There are two primary types of operators in lac operon:
Primary and Secondary
The primary operator is palindromic and positioned directly upstream from the promoter, whereas the secondary ones are placed on both sides of the primary one but with a much lower affinity to the repressor.
The three structural genes encodes enzymes necessary for lactose metabolism.
The first gene is lacZ, which encodes the β-galactosidase, an enzyme that cleaves lactose into glucose and galactose. However, β-galactosidase can also metabolize lactose to the allolactose, a metabolite which on interaction with repressor forces it to dissociate from the operator and thus turns on the operon.
The second gene is lacY that encodes the lactose permease, a membrane protein that facilitates the transport of lactose into the cell.
LacA encodes β-galactoside acetyltransferase, which transfers acetyl groups from acetyl-CoA to galactosides, detoxifying some compounds.
All three genes are transcribed together as a polycistronic mRNA, and the proteins are translated together to degrade lactose. The activity of the operon is controlled by the availability or non availability of lactose in the environment.
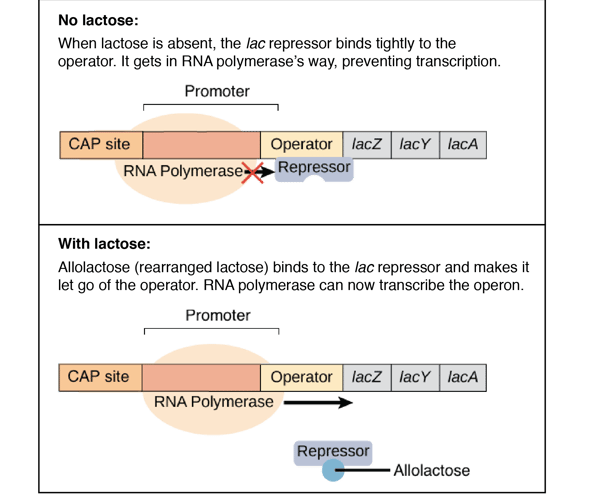
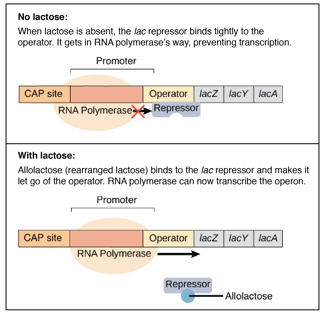
Negative Regulation of the Lac Operon: Regulation in the Absence of Inducer
How the Lac Repressor Inhibits Transcription in the Absence of Inducer
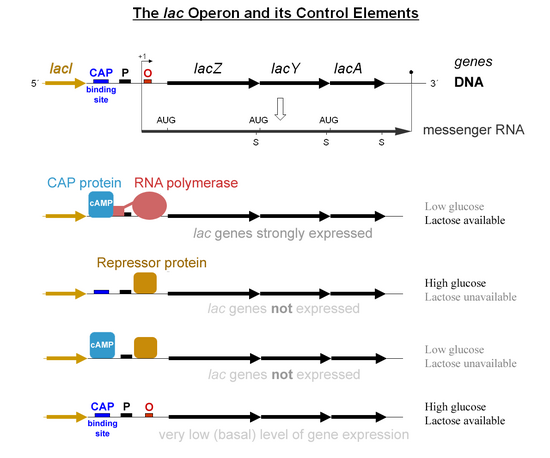
In the absence of an inducer, a product of the lacI gene, the lac repressor binds to the operator site of the lac operon and thereby inhibits transcription of structural genes. This is called negative regulation, which acts to keep genes from being expressed unless they are needed to ensure that energy is not wasted producing useless proteins.
This protein, coded by the lacI gene, is homotetrameric in structure and consists of four identical subunits weighing about 38,000 daltons. The repressor specifically binds to the operator sequence which consequently prevents the RNA polymerase from initiating transcription.
By binding to the operator, it would consequently prevent access of RNA polymerase to the promoter region; hence, transcription and the consequent expression of the proteins responsible for the metabolism of lactose is stopped.
Induction of the Lac Operon: Derepression by Allolactose
An inducer, allolactose, being one of a group of metabolites from lactose, when bound to the allosteric site of the repressor protein causes a conformational change that reduces the affinity of the repressor for the operator. It can thereby open up the promoter region for the binding of RNA polymerase and transcription.
Consequently, the lac operon is induced in the presence of lactose, and the necessary enzymes for the metabolism of lactose are made.
Isopropyl β-D-1-thiogalactopyranoside (IPTG), a synthetic non-metabolizable analog of lactose, also acts as an inducer. It is an example of a gratuitous inducer, a type of inducer that can activate the lac operon without being a substrate for β-galactosidase.
The expression of the lac operon in E. coli is an example of an inducible system. An inducible system is one that is activated in the presence of a substance known as an inducer. An inducer is a small molecule that triggers gene transcription by binding to a regulatory protein. In the lac operon, allolactose and its analogs that lead to the expression of the lac genes are called inducers, and the process of relieving repression on the operon is known as induction.
ADVERTISEMENTS
ADVERTISEMENTS
Positive Regulation and Catabolite Repression in Bacteria
How Bacteria Prioritize Carbon Sources in the Presence of Glucose
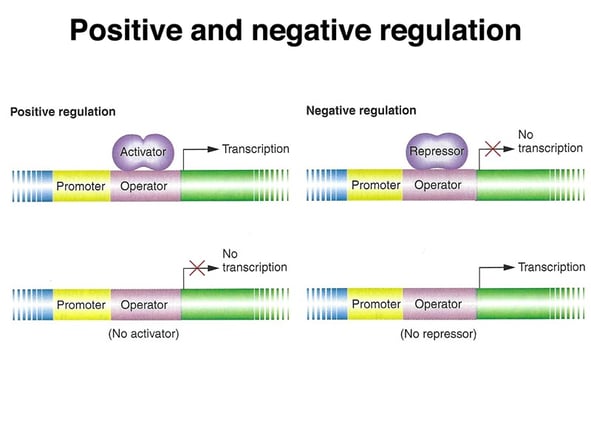
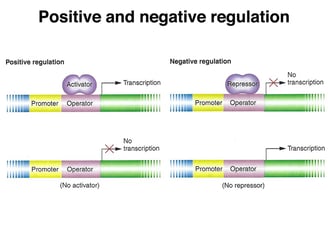
If a microbe is offered several carbon sources, the decision is made. In the presence of glucose and a secondary sugar, such as lactose, bacteria utilize glucose first and the use of the second sugar is inhibited. This is an example of catabolite repression, one type of regulatory system where one substrate inhibits its own use in the presence of another substrate.
Catabolite repression can be described as inhibition of certain enzymes which are involved in the catabolism of non-glucose sugars through glucose. Jacques Monod first discovered the phenomenon when he observed the following: bacteria preferentially catabolize glucose when a combination of glucose and another sugar is offered, causing them to exhibit a characteristic two-phase growth known as diauxic growth.
It later became known as "catabolite repression," where the presence of glucose repressed the biosynthesis of enzymes that participate in the utilization of non-glucose sugars for growth.
Catabolite repression is mediated by a protein called catabolite activator protein (CAP) that binds to a particular DNA sequence known as the CAP site. The activity of CAP is directly regulated by cAMP, whose production is inversely proportional to the concentration of glucose. In the presence of high glucose, cAMP is depressed and consequently CAP remains in an inactive status. When glucose levels become depressed, cAMP is generated and increases the concentration of active CAP, resulting in the production of enzymes capable of utilizing non-glucose sugars.
It is due to the regulatory mechanism of bacteria, which enables the bacteria to finely utilize available carbon sources as well as adapt to diverse environmental conditions.
Constitutive and Uninducible Lac Mutation
Understanding Lac Operon Mutations in E. coli
A mutant in which the structural genes of an operon are always transcribed, even in the absence of an inducer, is known as a constitutive mutant. In constitutive lac mutation, the lac operon genes are transcribed continuously—they cannot be turned off, even in the absence of lactose (i.e., the expression of the lac operon genes cannot be repressed). Constitutive mutations are more common in negatively regulated operons than in positively regulated ones.
On the other hand, an uninducible mutant is different. In uninducible mutants, the expression of lac operon genes cannot be induced. Lac mutations can also be categorized as dominant or recessive. To determine whether a particular lac mutation is dominant or recessive, a wild-type lac operon is introduced into a strain with the lac mutation.
For example, consider the lacI gene, which codes for the repressor. The wild-type (lacI⁺) and mutant (lacI⁻) forms differ in function. If the mutant lacI is recessive, introducing a normal lacI⁺ gene into the cell restores control, regardless of the presence of the mutant gene. Most lac mutations are recessive to wild-type and can be cis- or trans-acting. Recessive mutations that cannot be complemented are cis-acting, meaning they only affect the expression of genes on the same DNA strand where the mutation occurs.
Common Lac Mutations
1. lacI⁻ mutation: lacI⁻ mutants either fail to synthesize the repressor or produce a repressor incapable of binding to the operator. E. coli with lacI⁻ mutants exhibit constitutive expression of the lac operon. lacI⁻ mutants are generally recessive to the wild-type and are trans-acting.
2. lacI^s mutation: lacI^s mutants produce a repressor that cannot bind to the inducer, rendering the operon uninducible. The lacI^s mutation is dominant, trans-acting, and uninducible.
3. lacI^d mutation: This mutation arises because the Lac repressor protein forms a homotetramer, with monomers associating randomly in the cell. A mixture of normal and defective subunits can be non-functional, resulting in a constitutive phenotype similar to lacI⁻. These dominant-negative mutations occur in the DNA-binding site of the repressor polypeptide, making lacI^d dominant, trans-acting, and constitutive.
4. lacO^c mutation: lacO^c mutations are dominant, cis-acting, and constitutive. A mutation in the lac operator prevents the normal repressor from binding to the operator.
Expression of Lac Operon in Merozygotes
Certain strains of E. coli have the lac operon incorporated into an F' plasmid. These F' strains can pass the F plasmid to other F- strains, resulting in cells that are functionally diploid for the lac operon. A partially diploid bacterial cell, containing an exogenote donated by the F' plasmid, is known as a merozygote or partial diploid.
Jacob developed merozygotes for the lac operon genes to study their regulation. A culture of these diploid cells, which contain two sets of lac genes, is tested for regulatory phenotypes. This experiment, where genes or gene clusters are tested pairwise, is called a complementation test.
Case 1: Lac Operon with a lacO^c Mutation
In E. coli with a lacO^c mutation, the operator cannot bind the repressor, resulting in constitutive transcription even in the absence of lactose.
Merozygote State: If a bacterial chromosome contains a mutant operator (lacO^c) and the F' plasmid carries a wild-type operator (lacO⁺), the phenotype will be constitutive because the mutant operator exerts a cis-dominant effect. The wild-type operator on the plasmid cannot influence the chromosomal mutation.
Case 2: Lac Operon with a lacI^d Mutation
A lacI^d mutation produces a dominant-negative phenotype. In this case, the mutant repressor monomers interfere with the function of the normal repressor, resulting in constitutive transcription even in the presence of wild-type repressor genes.
Merozygote State: The presence of both wild-type and mutant repressor monomers in a merozygote leads to a constitutive phenotype since the mutant subunits spoil the function of the entire tetramer.
Case 3: Lac Operon with a lacI^s Mutation
In E. coli with a lacI^s mutation, the repressor cannot bind to the inducer, making the operon uninducible. Even in the presence of lactose, the lac operon remains repressed.
Merozygote State: Introducing a wild-type repressor gene on an F' plasmid in a cell with a lacI^s mutation can restore normal inducibility because the wild-type repressor compensates for the defective one.
ADVERTISEMENTS
ADVERTISEMENTS